

Studies in the Cohen lab span the entirety of spermatogenesis, asking a simple question: how are the unique events and transitions of spermatogenesis and oogenesis regulated at the genetic level to ensure the production of healthy viable gametes. We are particularly interested in the events of meiosis since it is during meiotic prophase I that numerous errors can arise that result in the production of gametes that are either unable to give rise to a healthy embry or that result in premature miscarriage or birth defects in humans. Indeed, some 40-60% of all human conceptuses are thought to have the wrong number of chromosomes due to errors in meiosis. Thus, while the lab studies virtually all aspects of spermatogenesis and oogenesis (see projects outlined below), we are particularly intrigued by meiosis.
In early meiotic prophase, SPO11 creates hundreds of DNA double-strand breaks (DSBs) throughout the genome. Spermatocytes and oocytes repair these DSBs via homologous recombination, producing either crossovers (sites of physical exchange of DNA between parental chromosome pairs) or noncrossovers. Crossovers serve two essential purposes: 1) Shuffling alleles into new combinations, facilitating genetic diversity in a population; and 2) Physically tethering homologous chromosomes together, ensuring their proper segregation into separate daughter cells. It is critical that at least one crossover forms between each chromosome pair as a safeguard against abnormal numbers of chromosomes (aneuploidy) in the resulting gametes – and ultimately offspring. The frequency, timing, and placement of crossovers are highly regulated and are sex- and species-specific. For instance, human and mouse males have significantly lower crossover numbers than females, and males preferentially form distally-placed crossovers. Although DSB number is sexually dimorphic in humans, the frequency is similar between male and female mice. In both species and sexes, only a small proportion of DSBs are resolved as crossovers (~10% in mouse; 28 - 37% in humans). Of the DSBs that form crossovers, a large majority (90-95%) are resolved via the class I crossover pathway, which utilizes MutS-gamma (MSH4/MSH5) and MutL-gamma (MLH1/MLH3). A second, minor class II crossover accounts for the remaining 5-10% of crossovers. In this latter pathway, the structure-specific nuclease MUS81 works in concert with BLM helicase to resolve complex multichromatid join molecule intermediates.
Since 2000, the Cohen Lab has studied meiotic recombination and the events of prophase I in mammals, and has been funded by NIH throughout this time. Our studies focus on the class I and class II crossover pathways, and their regulatory control/interactions.
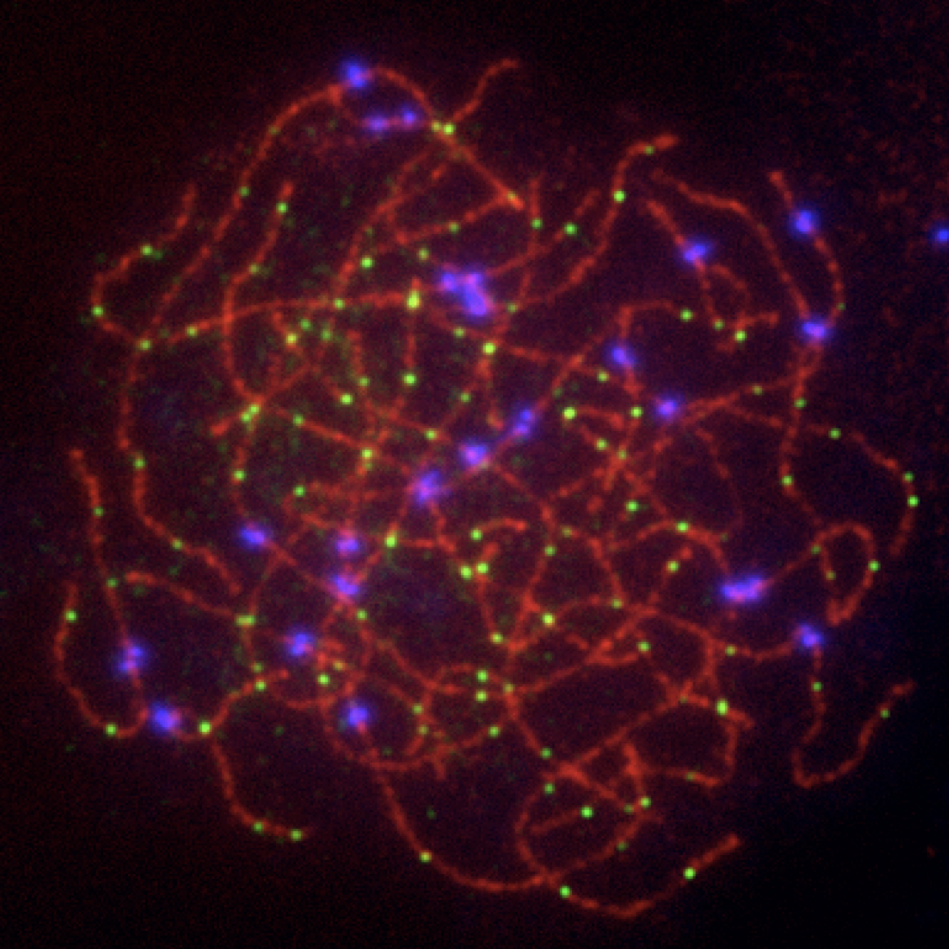
 MSH4-MSH5 rgulates crossover maturation in the mouse
MSH4-MSH5 rgulates crossover maturation in the mouse
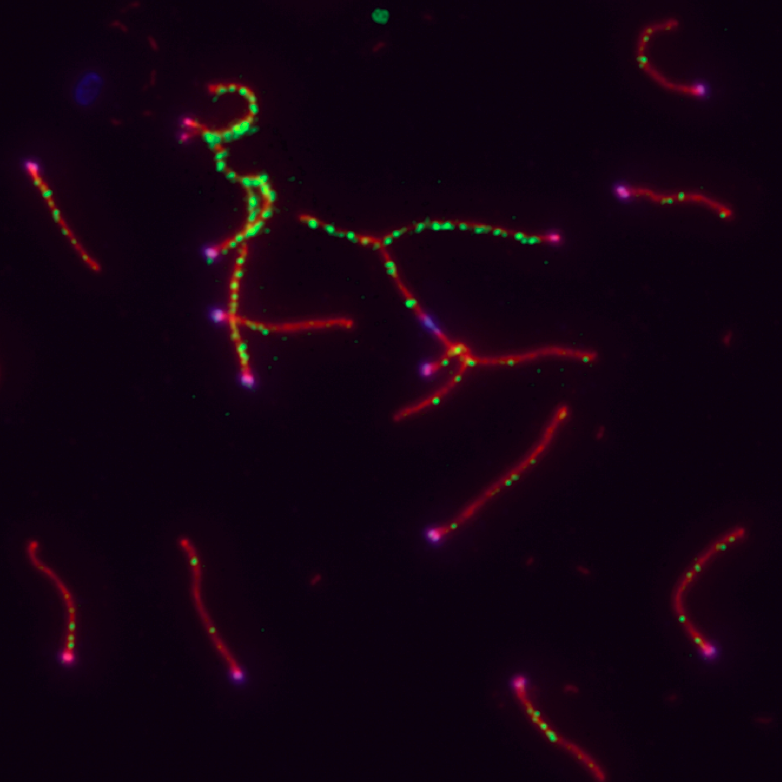
 The role of MLH1-MLH3 in driving class I crossovers in the mouse
The role of MLH1-MLH3 in driving class I crossovers in the mouse
 The role of CNTD1 in directing class I crossovers
The role of CNTD1 in directing class I crossovers
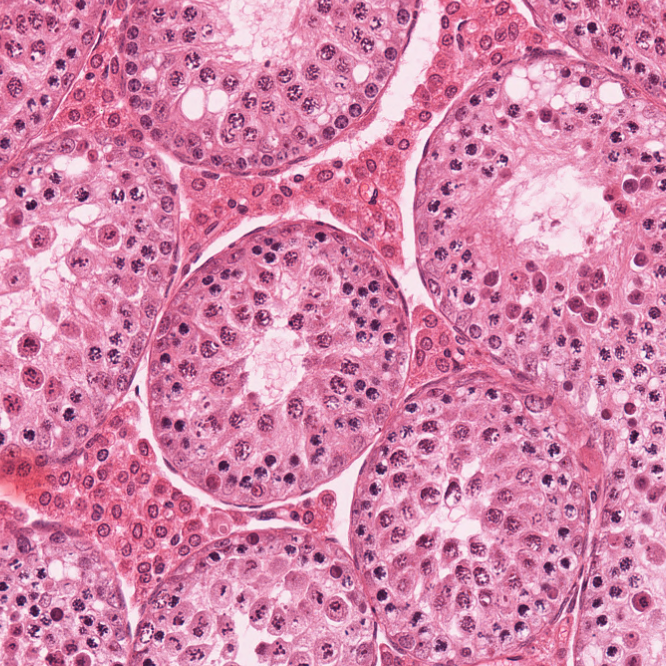
Our studies focus on the DNA mismatch repair pathway, the MUS81-EME1 structure specific nuclease, and the Fanconi Anemia pathway. We study these pathways in the context of meiotic recombination as well as in the context of genome stability in the mammalian germ line.
 The role of the MUS81-EME1 endonuclease in driving crossing over during meiosis
The role of the MUS81-EME1 endonuclease in driving crossing over during meiosis
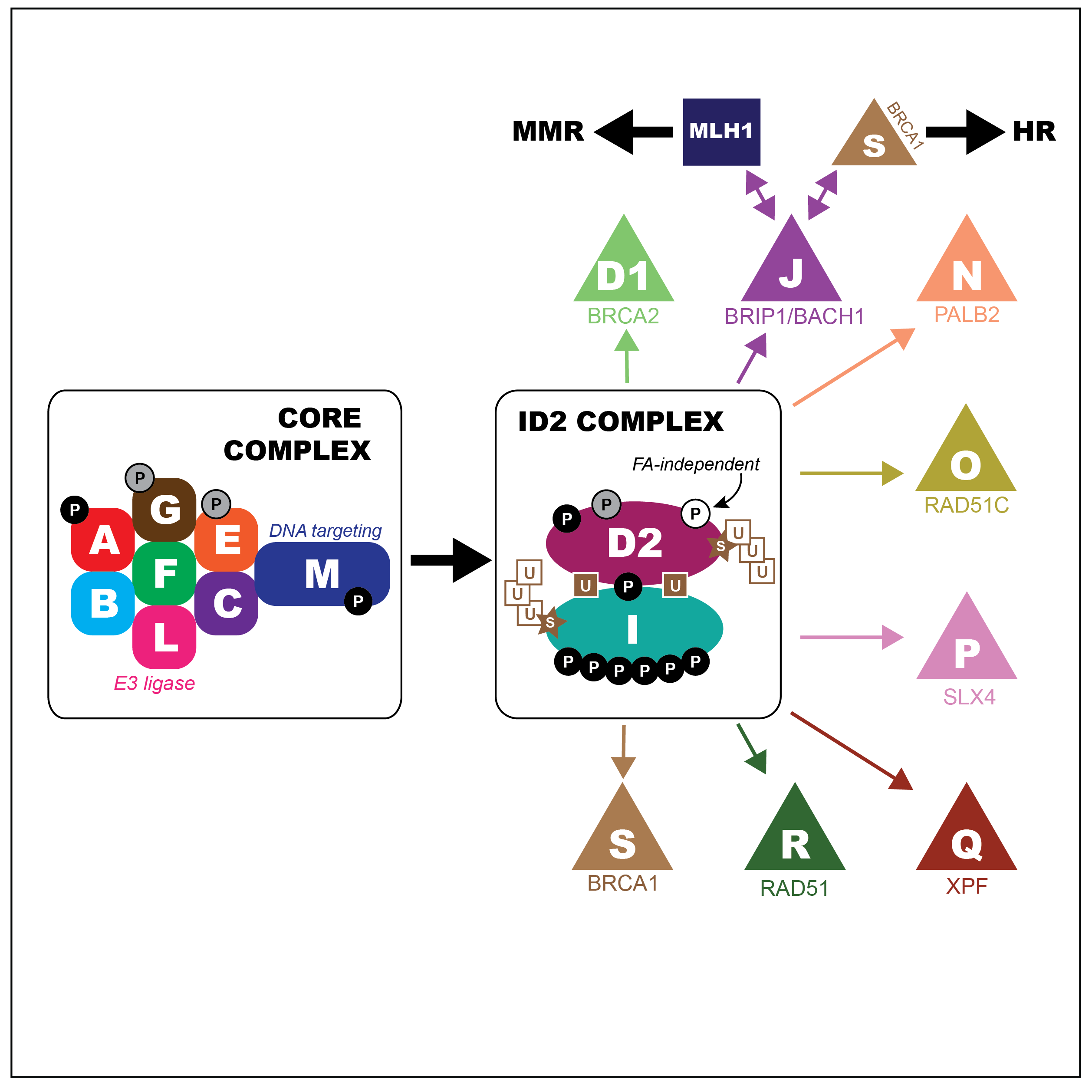 The role of the Fanconi Anemia pathway in gametogenesis
The role of the Fanconi Anemia pathway in gametogenesis
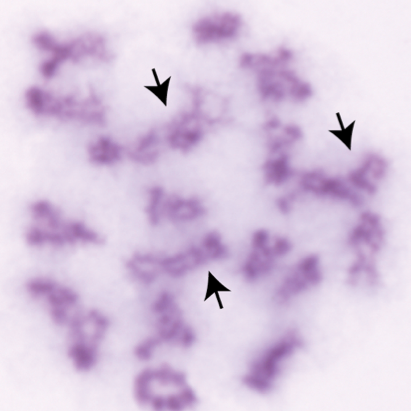 The role of BLM helicase in crossover decisions
The role of BLM helicase in crossover decisions
We seek to understand the complex gene regulatory environment that exists throughout spermatogeneis, from transcriptional regulation, to the role of non-coding RNAs in post-transcriptional regulation, and to the post-translational regulation of protein function.
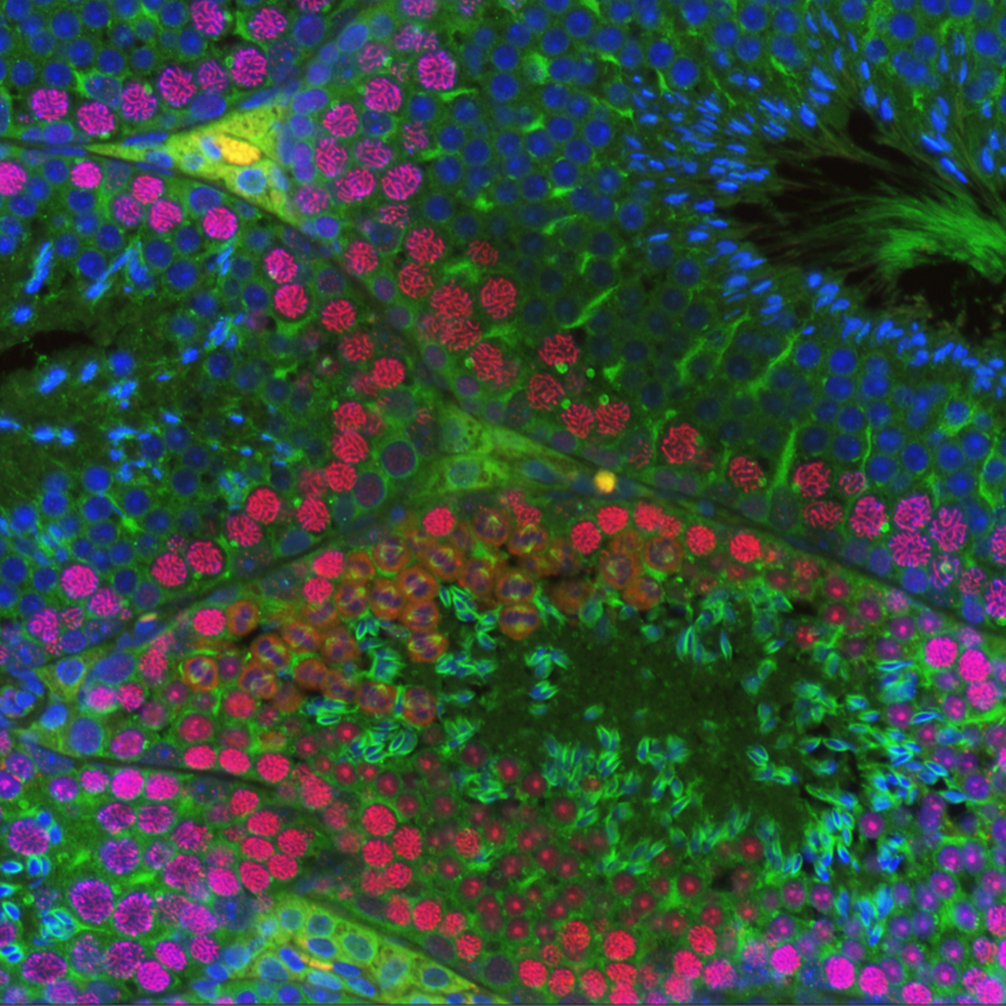 Regulation of gene expression throughout spermatogenesis
Regulation of gene expression throughout spermatogenesis
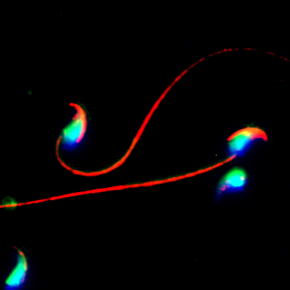 The role of non-coding RNAs in regulating spermatogenesis
The role of non-coding RNAs in regulating spermatogenesis
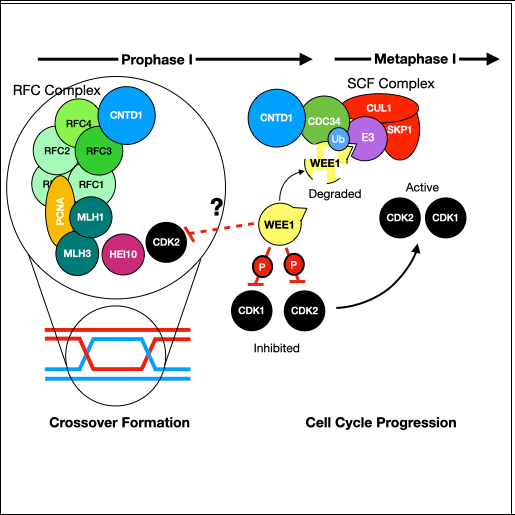 Post-translational control of protein function
Post-translational control of protein function
Studies in the lab are focused on identifying intervention points through spermatogenesis that could be targets of contraceptive drug targeting. Our goal is to provide cheap, safe, and reliable contraceptive options for men. Funded by the Bill and Melinda Gates Foundation.
 Investigating novel strategies for contraception
Investigating novel strategies for contraception
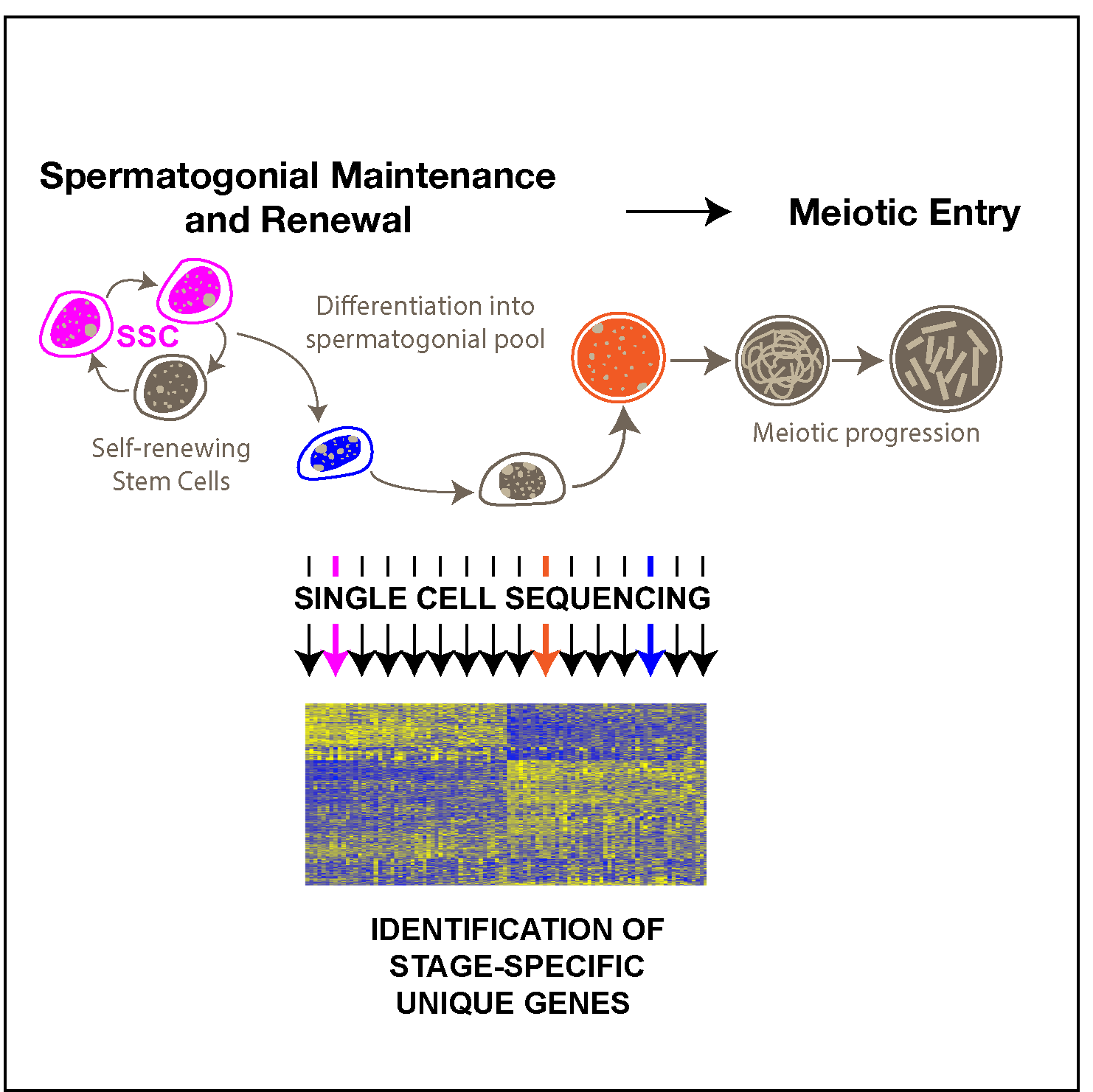 The genetics of meiotic entry
The genetics of meiotic entry